INTRODUCTION
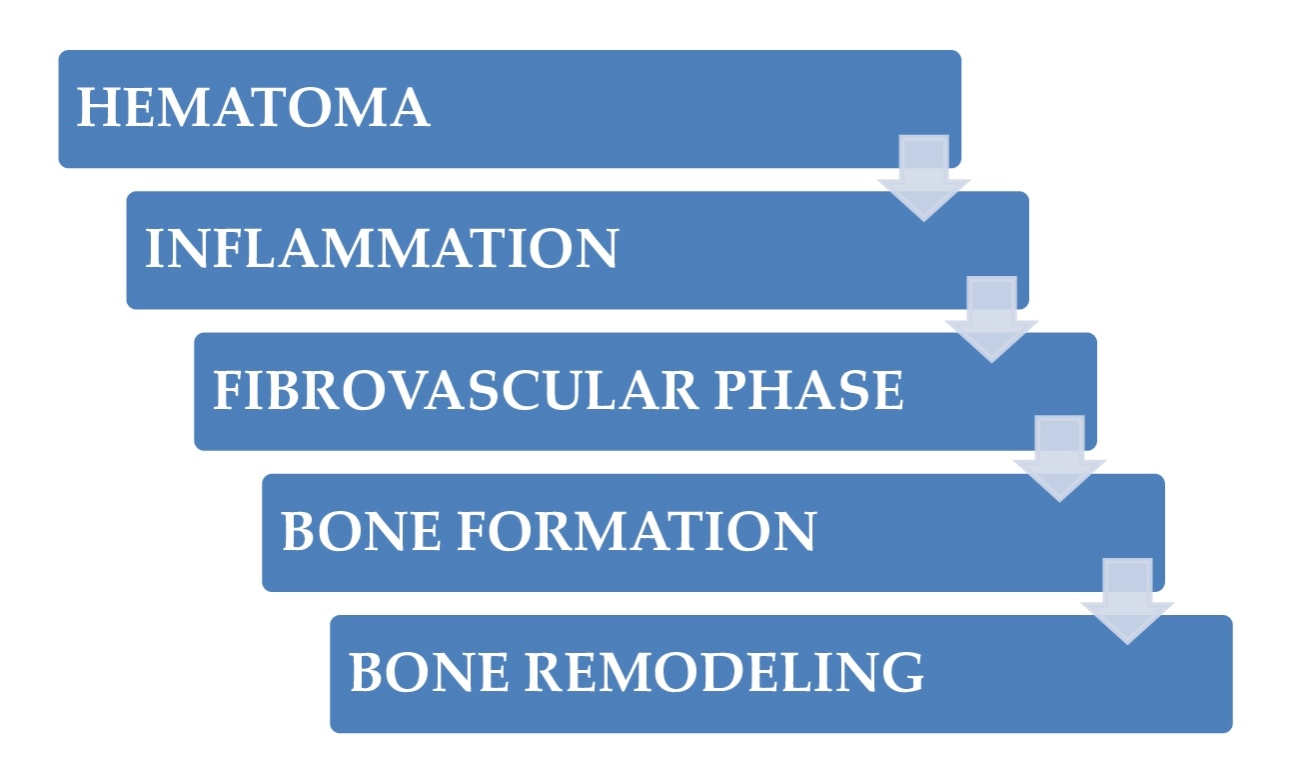
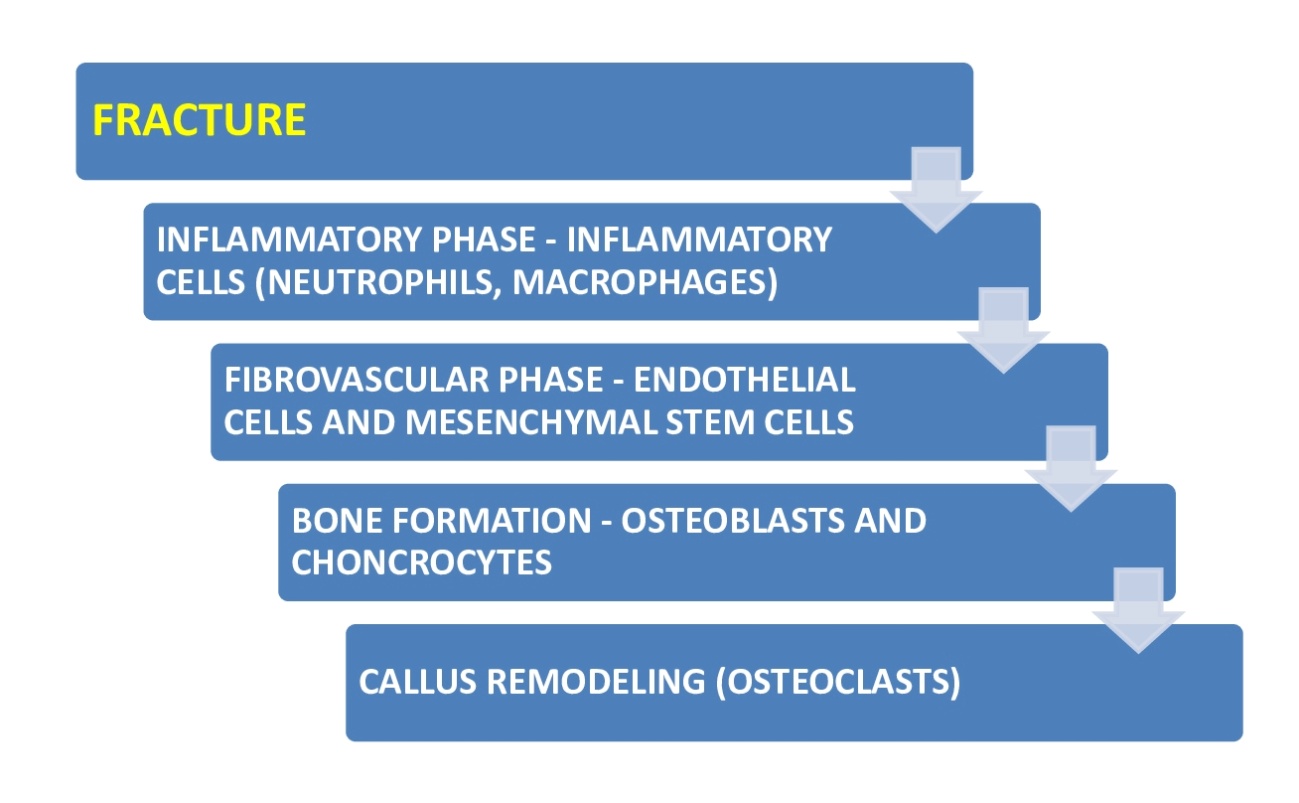
Bone fractures heal through two different mechanisms: Direct (primary) or indirect (secondary) healing.1 Primary healing entails a direct shift of mesenchymal stem cells (MSCs) to bone-forming osteoblasts (intramembranous ossification). Secondary healing moves forward through a cartilage intermediate before a bone is created by osteoblasts (endochondral ossification).1,2 The cellular and molecular elements coordinating fracture callus creation and resolution are intricate and incredibly arranged. This article will mainly review secondary healing since most fractures clinically heal in this mode. The mechanism of bone healing has a diversity of cellular constituents needed for the advancement of healing. Inflammatory cells (i.e., mast cells, macrophages, and neutrophils) are the first cellular constituent of the fracture ambiance, followed by MSCs, endothelial cells, chondrocytes, osteoblasts, and osteoclasts. The mechanism of fracture healing can be deemed in episodic temporal sections; however, it is essential to know that there is a substantial overhang of the temporal sections of healing, and associated cell categories exist together. This is a fundamental notion because cell-to-cell signaling, in a heterotypic way (across cell types), is beyond question crucial.1–3 Figures 1 and 2 summarize the main phases of bone healing and the cells implicated in each.


This article aimed to perform a narrative review of the literature on recent advances concerning cells involved in bone healing and their possible clinical applications. On December 26, 2022, a PubMed literature search used “cell bone repair” as a keyword. A total of 26,145 articles were found, of which 74 were analyzed that met our inclusion criteria.
INFLAMMATORY PHASE – INFLAMMATORY CELLS
This phase has three main parts: acute inflammation, resolving inflammation, and chronic inflammation.
Acute Inflammation
Inflammatory cells are deposited along the clot during hemorrhage, moving to the fracture site from local sources. One function of inflammatory cells, specifically neutrophils, macrophages, and mast cells, is the debridement of injured and devitalized tissue. Inflammatory cells also create cytokines that positively and negatively affect healing.4–6 Some of these cytokines are found at the fracture site within the first 24 hours post-injury and are essential for expanding the inflammatory reaction by acting on cells in the bone marrow, periosteum, and hematoma.7,8 Macrophages produce the pro-inflammatory molecule interleukin-1 (IL-1). IL-1 controls the expression of cyclooxygenases (COX-1 and COX-2), which are the enzymes that form prostaglandins in the fracture site.9 In addition to rendering inflammatory cytokines, inflammatory cells also create growth factors such as fibroblast growth factors (FGFs), platelet-derived growth factor (PDGF), and transforming growth factor-beta (TGF-beta), which start the repair mechanism by promoting proliferation and differentiation of the MSCs that give rise to the fracture callus.10–12
Recent advances in neutrophils
Zhang et al. found that in reaction to fracture injury, the percentages of neutrophils and associated plasma extracellular vesicles (EVs) were substantially greater in fracture calluses than in peripheral blood.13
Recent advances in macrophages
It has been shown that dexamethasone diminishes the amount of macrophages at the injured area during early osseous repair following femoral bone injury partially through plasminogen activator inhibitor-1 (PAI-1) and macrophage colony-stimulating factor (M-CSF) in mice.14 Kohara et al. have reported the relevance of urokinase plasminogen activator (uPA) and transforming growth factor beta 1 (TGF-β1) during osseous regeneration, showing a new process of osseous regeneration mediated by macrophages.15
Toita et al. has exposed the pros of local M1-to-M2 macrophage polarization prompted by phosphatidylserine-containing liposomes (PSL)-multilayers constructed on implants for efficacious osseous regeneration and osseointegration (bone-to-implant integration).16 It has been observed that macrophage recruitment scarcity in nearby soft tissue in early surgery for high-energy fractures could be a significant cause of atrophic nonunion.17 Baratchart et al. reported the following essential data: antiinflammatory macrophages are crucial for early osteoclast constraint and proinflammatory macrophage suppression; proinflammatory macrophages are implicated in osteoclast osseous resorptive activity, but osteoblasts facilitate osteoclast differentiation.18
Recent advances in mast cells
It has been recently encountered that mast cells participate in trauma-caused impaired bone repair. It could be a possible objective for new treatment alternatives to facilitate fracture healing in multiple injured individuals.19 Fischer et al. have shown that mast cells adversely impact the healing of bone fractures under estrogen-deficient conditions. Targeting mast cells could be a new therapeutic strategy to ameliorate impaired bone repair in postmenopausal osteoporosis.20
Resolving inflammation
While the inflammatory stage of fracture healing starts during the earliest phases of repair, it has been shown that the inflammatory cells are also present throughout later stages and seem to experience changes as healing continues.21 Studies on fracture healing in mice lacking the tumor necrosis factor-alpha (TNF-alpha) receptor showed delays during acute inflammation and later phases of healing.22,23 Additionally, IL-6 expression seems bimodal during fracture healing, indicating a temporally specific function for inflammatory cytokines during bone repair.24
Chronic Inflammation
Chronic, non-resolving inflammation is harmful to fracture healing. Experimental data have shown that fracture healing and osseointegration are disjointed in diseases where there is chronic, non-resolving inflammation, such as diabetes.25–27
FIBROVASCULAR PHASE – ENDOTHELIAL CELLS AND MSCs
Following inflammation, the angio-mesenchymal stage of repair starts. This stage has been called the fibrovascular stage. It is characterized by vascular remodeling (angiogenesis and neovascularization) and recruitment of MSCs that eventually differentiate into chondrocytes and osteoblasts to regenerate the fractured bone.
Revascularization
During the initial fracture trauma, vascular supply is reinstituted quickly by developing a new vascular network.28 Creation of the network happens by two distinct mechanisms: Angiogenesis and vasculogenesis. Angiogenesis is how new blood vessels are created by blooming from existing vasculature. Vasculogenesis is the de novo creation of blood vessels from in situ endothelial progenitor cells (EPCs) within the callus. Endothelial cells forming callus vasculature can develop from several sources, including existing vessels of the periosteum and the intramedullary vasculature,29 circulating EPCs augmented during fracture repair,30,31or the bone marrow.32
Vascular endothelial growth factor (VEGF) is a well-characterized driver of angiogenesis and vasculogenesis.33 VEGF is produced by several cells in the fracture callus, including inflammatory cells, mesenchyme, osteoblasts, and hypertrophic chondrocytes. VEGF binds the VEGF family of receptors VEGFR1 (FLT1) and VEGFR2 (FLK1), activating signaling cascades that result in augmented proliferation and blooming of endothelial cells and recruitment of EPCs to the fracture.34
Endothelial cells and MSCs
Most MSCs recruited to the fracture site come from the local periosteum and bone marrow. Recruitment of MSCs in the fracture repair process is under molecular control by cytokines released at the fracture site, especially CXCL12, also known as stromal cell-derived factor 1 (SDF1). The damaged periosteum liberates SDF1 and drives mobilization and homing of MSCs through CXCR4.31 Notch signaling is another possibly crucial factor in controlling MSC amount and activation.35
Recent advances in endothelial cells
He et al. have suggested that MSCs migrate toward endothelial cells (ECs) via PDGF-BB/PDGFRβ and the downstream Src-Akt signal pathway in the inflammatory microenvironment.36
Recent advances in MSCs
Esposito et al. have identified a new population of endosteal cells that is functionally controlled through the modulation of CXCR4 by IGF-1R signaling, and such control is crucial in osseous homeostasis and fracture healing. This knowledge could help in the development of new therapeutic methods by targeting CXCR4 signaling to manage nonunions.37
According to Jeffery et al., distinct osseous injuries are repaired by particular skeletal stem/progenitor cells (SSCs), with periosteal cells regenerating bone and marrow stroma following non-stabilized fractures.38 TABLE 1 summarizes recent advances in MSCs.39–47
BONE FORMATION – OSTEOBLASTS AND CHONDROCYTES
Following the fibrovascular stage of healing, many of the MSCs that formed the fibrovascular callus experience differentiation to either osteoblasts or chondrocytes to start the bone formation stage of healing. Differentiation of MSCs into bi-potential osteochondral progenitor cells is initially regulated by Sox9.48 Factors controlling the choice of progenitor cells towards the chondrogenic or osteogenic destiny are multifactorial, integrated, and still being established. Extrinsically, mechanical factors and oxygen tension are crucial variables in fate decisions.49,50 The aforementioned cell-extrinsic factors lead to particular cell-intrinsic regulation of chondrogenesis and osteoblastogenesis. Secreted growth factors also have a direct impact on MSCs differentiation. Bone morphogenetic proteins (BMPs) are the classic osteogenic molecules related to bone formation. The Wnt family is another secreted growth factor family that could play a role in controlling MSCs fate determination in bone healing.3
Intramembranous ossification - osteoblasts
Intramembranous bone creation from these endosteal stem cells is responsible for quickly bridging across the marrow cavity.51
Recent advances in osteoblasts
Type 2 diabetes T2D had adverse effects on bone healing via inhibition of osteoblast differentiation of skeletal stem cells and induction of hastened bone senescence, and that the hyperglycemia per se and not just insulin levels was detrimental for bone healing.52 Jing et al. have stated that despite some debate concerning the function of signaling pathways in osseous creation, the Wnt/β-catenin, Notch, PI3K/Akt/mTOR, Runx2, IGF, FGF, and BMP/TGF-β cascades contribute to the efficacy of osseous regeneration by augmenting osteogenesis and maturation of osteoblasts.53 A study has demonstrated that intraflagellar transport 140 (IFT140) benefits fracture repair.54 Ahmad et al. have suggested that Cdk5 controls osteoblast differentiation through MAPK pathway modulation. They stated that Cdk5 is a possible therapeutic objective to manage osteoporosis and ameliorate fracture healing.55
Endochondral bone formation - chondrocytes
Conversion of the cartilage callus to bone follows a highly regulated maturation of chondrocytes from a proliferative through a hypertrophic state.56 Chondrocyte hypertrophy is an essential state during endochondral ossification. Hypertrophic chondrocytes are highly angiogenic and promote a second phase of vascular invasion into the cartilage callus by synthesizing VEGF,57–59 PDGF,60 and PlGF (placental growth factor).61 The molecular stimulus for calcification is not fully understood, but BMPs probably play a vital role in this process. BMP is expressed by both hypertrophic chondrocytes,62 and vascular endothelial cells,63 indicating that BMP signaling has both cell-autonomous and paracrine effects that might drive calcification. After calcification of the cartilage, bone formation happens. Histological staining in this vascularized transition area between cartilage and bone shows hypertrophic chondrocytes entrapped in a bone matrix next to the vasculature.64 As the cartilaginous matrix disappears and a bone matrix is laid down, the large round hypertrophic morphology of chondrocytes is gradually converted into morphology characteristic of the osteocytes with cellular extensions existing in canaliculi.56,62–64 Hypertrophic chondrocytes can become osteoprogenitors and osteoblasts, directly contributing to woven osseous creation.65
Bai et al. identified a long non-coding RNA (lncRNA) named hypertrophic chondrocyte Angiogenesis related lncRNA (HCAR). They demonstrated it to facilitate the endochondral osseous repair by upregulating the expression of matrix metallopeptidase 13 (Mmp13) and vascular endothelial growth factor alpha (VEGF-alpha) in hypertrophic chondrocytes. Lnc-HCAR knockdown in hypertrophic chondrocytes inhibited the cartilage matrix remodeling and diminished the CD31hiEmcnhi vessels amount in an osseous repair model.66 Stegen et al. have reported a crucial role of phosphoglycerate dehydrogenase (PHGDH)-dependent serine synthesis in preserving intracellular serine levels under physiological and serine-limited conditions, as appropriate serine levels are required to support chondrocyte proliferation throughout endochondral ossification.67
CALLUS REMODELING AND OSTEOCLASTS
A critical component of callus remodeling is bone degradation by osteoclasts.64 Osteoclasts originate from hematopoietic monocyte/macrophage lineage precursors. Proliferation and survival of osteoclast precursors is triggered by interaction between monocyte / M-CSF) and its receptor c-fms, which is present in both macrophages and osteoclasts. Bone marrow macrophages differentiate into osteoclasts upon stimulation with the Receptor Activator of Nuclear Factor kappaB Ligand (RANKL), which binds to its receptor, RANK. Osteoclast differentiation happens through multiple stages.64,68 Both MCSF and RANKL are needed throughout the differentiation process and contribute to mature osteoclasts’ survival. MCSF and RANKL are both required and sufficient for osteoclast creation and function, but multiple other cytokines and signaling pathways influence osteoclast differentiation, maturation, and survival.69–72
Recent advances in osteoclasts
Wu et al. have reported that small extracellular vesicles (sEV)-mediated miR-106a-5p transfer plays a crucial function in osteogenesis and indicates a new communication strategy between osteoclasts and bone MSCs (BMSCs).73 BM-MSCs -based and macrophage-based cell treatment are regarded as hopeful approaches to facilitate fracture healing because of the amazing osteogenic potential of BM-MSCs and the typical immunomodulatory role of macrophages. Besides, evidence has also shown the importance of cross-talk between these two cell types in the fracture healing process.74
CONCLUSIONS
The following conclusions can be drawn from the literature review:
-
Cross-talk between MSCs and macrophages is crucial in fracture healing.
-
Rejuvenation of neutrophils and their EVs is associated with improved aged fracture healing.
-
A new population of endosteal cells that are functionally controlled through the modulation of CXCR4 by IGF-1R signaling has been identified, and such control is crucial in osseous homeostasis and fracture healing. This information could help to develop new therapeutic methods by targeting CXCR4 signaling to manage nonunions.
-
BM-MSCs-EVs carrying miR-29b-3p promote fracture healing.
-
HUCMSCsWnt10b might be a promising treatment for long bone fractures.
-
Aucubin facilitates bone-fracture healing by partially controlling the osteogenesis of hBM-MSCs via the canonical BMP-2 / Smads signaling pathway and by suppressing oxidative stress damage partially via the Nrf2/HO1 signaling pathway.
-
Early treatment of fractures with bortezomib could improve fracture repair by augmenting the amount and proliferation of MSCs.
-
Episodic alcohol exposure could affect normal fracture healing through the mitigation of MSCs chondrogenic differentiation at the callus site.
-
Dexamethasone diminishes the amount of macrophages at the injured zone during early osseous repair.
Forthcoming investigations on the value in clinical practice of Aucubin, Bortezomib, and human umbilical cord MSCs (HUCMSCs)Wnt10b to improve bone healing in managing bone fractures are crucial. Future guidelines should consider that episodic alcohol exposure and Dexamethasone treatment may affect bone healing if employed in individuals who are being treated for a bone fracture.
DECLARATION OF CONFLICT OF INTEREST
The author does NOT have any potential conflicts of interest for this manuscript.
DECLARATION OF FUNDING
The author received NO financial support for the preparation, research, authorship, and publication of this manuscript.
DECLARATION OF ETHICAL APPROVAL FOR STUDY
There is no approval required for review papers at this institution.
DECLARATION OF INFORMED CONSENT
There is no identifiable information in this submission.